
生命史理论概述及其与社会心理学的结合之以道德行为为例
分类:文史论文
时间:2022-02-23
摘 要 生命史理论作为进化心理学的代表性理论之一, 同时关注了环境因素和个体经验, 是社会心理与个人心理的有机结合, 但是该理论目前在社会心理学领域得到的关注还相当有限。因此该理论具体的建立、发展过程, 主要理论观点, 以及主要测量方法、测量工具都需要进行详细的梳理和介绍, 以便于研究者对其有较为全面的理解。此外, 结合社会心理学的重要研究主题——道德行为, 可以发现不同生命史策略在个体的亲社会行为、不道德/暴力犯罪行为上可能有不同的表现, 并且控制感、安全感和调节定向理论都会是其可能的影响机制。因此在未来的研究中, 应该将生命史理论在社会心理学领域进行更广泛的应用, 为社会心理学研究者提供一个分析道德行为的新视角。
关键词 生命史; 道德行为; 权衡; 影响因素; 测量方法
1 前言
近年来, 随着进化心理学的发展, 传统心理学家对该领域的关注不断增加(Chang, 2007), 而采用进化心理学理论对个体行为进行解释的研究也越来越多。生命史理论(Life History Theory, Kaplan & Gangestad, 2005; 也有译作生活史理论) 就是进化心理学中具有代表性的理论之一。该理论认为, 个体在资源有限的情况下, 需要考虑如何分配自身资源, 而个体所处环境、所经历的生活事件则会影响个体形成不同的分配策略(即生命史策略)。
虽然生命史理论同时关注了环境因素和个体经验, 是社会心理与个人心理的有机结合, 可以拓展到社会心理学领域的研究中, 但是该理论在社会心理学领域的应用还相当有限。生命史理论主要关注个体对资源的分配, 而在社会心理学领域与资源相关的一个重要的研究领域就是道德行为。例如个体将资源与他人共享属于亲社会行为, 而牺牲他人利益来获取资源则属于不道德行为 (Figueredo, Adnrzejczak, Jones, Smith-Castro, & Montero, 2011; Wiebe, 2012)。虽然生命史理论并未直接预测环境和过往经历会如何影响个体的道德行为, 但是基于以往研究, 二者具有紧密联系。因此本文就以道德行为为例, 尝试将生命史理论应用于社会心理学研究中去。
要对生命史理论进行扩展和应用, 首先需要对该理论有详细的了解, 鉴于以往并没有文献体系地介绍过该理论, 本文首先介绍了生命史理论的建立、发展过程和主要理论观点, 包括生命史理论中基本的权衡问题、快慢生命史策略的概念、影响生命史策略的因素以及主要测量方法; 接着整合相关研究成果, 将生命史对个体行为影响的效应扩展至社会心理学领域中, 探索不同的生命史策略如何体现在个体的道德行为上(具体为亲社会行为和不道德/暴力犯罪行为), 期望能从新的角度更整体、更全面的了解生命史理论在社会心理学的应用, 扩展生命史理论的研究范畴。
2 生命史理论概述
繁衍和死亡是个体在生命进程中要面对的两个重要命题(Kaplan & Gangestad, 2005)。由于资源的有限性, 所有个体在生命进程中随时会面对如何最优化地分配资源以获得最大发展的问题 (Griskevicius, Tybur, Delton, & Robertson, 2011)。生命史理论正是解释所有有机体(包括人类), 如何形成和采取不同的生命史策略(Life History Strategy; Kaplan & Gangestad, 2005), 来对有限资源进行分配的过程(Charnov, 1993; Daan & Tinbergen, 1997; Low, 2000; Roff, 1992; Stearns, 1992)。
2.1 个体分配资源时的四类权衡
不同的生命史策略体现在个体面对一系列生命命题时, 如何权衡资源的分配。最初进化学家 MacArthur 认为, 个体分配资源时主要是在生存投入(Somatic Effort)与繁殖投入(Reproductive Effort) 间进行权衡(MacArthur & Wilson, 1967)。前者以生存为导向(Griskevicius, Tybur et al., 2011), 而后者则是指个体将资源主要分配于繁衍、养育后代等方面。而繁殖投入又进一步包括求偶权衡与养育后代这两方面的权衡(MacArthur & Wilson, 1967)。
后来 Kaplan 和 Gangestad (2005)在论述生命史理论时, 将个体分配资源时的权衡总结为三类:即“现在–未来繁殖权衡” (Present-future reproduction trade-off), “求偶–养育权衡” (Trade-off between mating effort and parenting effort), 以及“后代数量– 质量权衡” (Quantity-quality of offspring trade-off)。然而 Ellis, Figueredo, Brumbach 和 Schlomer (2009) 进一步提出, 在现在–未来繁殖权衡之前, 个体首先需要在维系与成长之间进行权衡(Trade-off between maintenance and growth), 它是个体进行“现在–未来繁殖权衡”的基础和平台。通过对相关文献的梳理, 研究者发现上述这四类主要的权衡问题是相互包含的, 并对其之间的关系进行了整理, 如表 1 所示。
维系–成长的权衡。个体在发展过程中面对的最基本问题是维系与成长之间的权衡。为了生存, 人们需要将所有精力与资源用来维系现有状况以抵御来自死亡的威胁; 当维系或者生存的需求获得基本满足后, 个体才会将资源更多用于成长, 并最终用于繁殖。
现在–未来繁殖的权衡。在解决了维系–成长的权衡问题之后, 个体则需要进一步权衡是在把资源继续用来发展自己还是留给后代, 因此也产生了何时繁衍的问题。“现在–未来繁殖权衡”是指个体在自我发展和繁衍后代之间的权衡, 如果个体选择现在繁衍后代, 就会耗费本可用于自身成长的资源, 必然会在一定程度上阻碍自己的发展; 而选择未来繁衍, 则可以满足当下的成长与维系之需, 但同样会产生风险, 比如不能获得当下繁衍可能带来的收益(Kaplan & Gangestad, 2005)。
求偶–养育权衡。求偶–养育权衡隶属于现在繁殖策略。将资源投入于求偶可以提高后代数量, 而投入于养育则提高后代的质量, 个体在这两个权衡间如何选择取决于在当下环境中哪种行为带来的收益更大。
后代数量–质量权衡。后代数量–质量权衡也隶属于现在繁殖策略, 由于养育者的资源有限, 后代数量增加时所消耗的资源必然导致每个后代所能获得的资源减少, 后代质量下降等结果, 因此个体在分配资源时需要权衡要增加后代的数量还是质量(Kaplan & Gangestad, 2005)。
2.2 “快”与“慢”的生命史策略
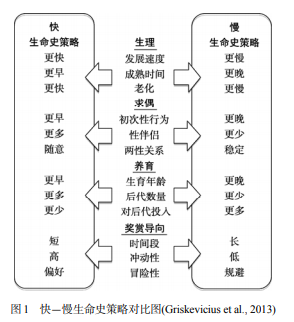
个体如何在上述四类权衡中选择, 就反映出个体不同的生命史策略。作为生命史理论中的重要概念, 生命史策略是个体特有的行为模式的集合, 个体独特的生活经历(即生命史)会影响个体的行为模式, 形成不同的生命史策略, 而不同的生命史策略所代表的行为也反映出了个体独特的生活经历。为了系统地描述生命史策略, Ellis 等人 (2009)进一步将各类生命史策略放在一个由“慢” 至“快”的连续体上进行评价(Fast-slow Strategy)。
具体来说, 慢生命史策略往往指向未来的生存投入(Somatic Effort), 比如更晚的生育年龄以及延迟满足行为(Figueredo et al., 2006), 而快生命史策略则指向当下的繁殖投入(Reproductive Effort), 如更早地生育, 更看重当下获利等, 具体如图 1 所示。
2.3 影响生命史策略的因素
不同的个体可能会持有不同的生命史策略, 而哪些因素会影响个体形成不同的生命史策略?总结近些年的相关研究结果, 研究者发现主要有三类影响生命史策略的因素, 包括环境因素、童年经历和生物因素。
2.3.1 环境因素
Ellis 等人(2009)认为环境的严酷性(Harshness) 和不稳定性(Unpredictability)是影响个体生命史策略的两个关键因素。其中环境的严酷性是指恶劣的外部环境对种群的破坏程度, 如资源匮乏 (Resource scarcity); 而环境的不稳定性则指环境的严酷性在时间和空间两个维度上的变化情况 (Ellis et al., 2009), 如环境的动荡(Environmental fluctuations)。这两种关键的环境因素可以通过社会中的疾病-死亡率(Mobility-Mortality)反映出来, 疾病–死亡率越高(high level of mobility-mortality), 说明环境越严酷, 而疾病–死亡率波动越大(high variation of mobility-mortality), 则说明环境越不稳定(Ellis et al., 2009)。由于人类的繁衍本能, 严酷、不稳定的环境往往会促使人类形成较快的生命史策略, 将更多资源投入于繁衍后代(Griskevicius, Delton et al., 2011)。
2.3.2 童年经历
除了环境因素外, 进化心理学家认为, 童年经历也会影响个体的生命史策略。Griskevicius 等人(2013)提出了敏感化模型(Sensitization Model) 来解释童年经历如何影响生命史策略的形成, 即不同的早期生活经验可以塑造人们成年后面对逆境时不同的应对方式。此外, Ellis 等人(2009)也认为童年经验可以使个体建构出对世界的普遍认识观, 这种认知图式影响个体形成不同的生命史策略。
Brumbach, Figueredo 和 Ellis (2009)的研究发现, 不论童年时期是暴露在极端恶劣或是不稳定的生活环境, 都会促使个体在成年早期形成更快的生命史策略(Ellis et al., 2009; Raley & Wildsmith, 2004)。而快的生命史策略可以加快儿童的生理发展 (Belsky, Houts, & Fearson, 2010; Ellis, 2004), 导致儿童过早的性成熟, 使月经初潮更早, 性生活更放纵(Figueredo et al., 2005), 以及更早开始生育(Griskevicius, Delton, Robertson, & Tybur, 2011)。
此外, 居住环境的变化也会使童年生活变得不稳定(Frankenhuis, Gergely, & Watson, 2013)。研究表明, 青少年时期频繁搬家与其快生命史策略呈高相关, 频繁搬家的个体可能初次性行为年龄更小, 在青春期倾向有多个性伴侣, 婚前性行为更普遍, 怀孕和成为单亲妈妈机率更大(South, Haynie, & Bose, 2005; Crowder & Teachman, 2004; Baumer & South, 2001)。
另外, 敏感化模型还提出, 个体的生命史策略只有在困难时期才会被激发出来, 在顺境中即使个体的童年经验不同, 他们的应对方式也无显著差异。例如 Griskevicius 和 Tybur 等人(2011)研究发现, 环境中有关死亡的线索可以影响个体的冒险倾向与延迟满足行为, 但这一影响会受童年时期家庭的社会经济地位的调节, 对于童年家庭环境贫困的个体, 给他们呈现暗示死亡的环境线索会促使他们采取快的生命史策略, 更愿意冒险去获得当下的利益; 而对于成长环境较富裕的个体, 死亡线索会促使他们采取更慢的生命史策略, 更看重未来的生活 , 并拒绝冒险。 White, Li, Griskevicius, Neuberg 和 Kenrick (2013)的研究也发现, 死亡威胁线索与童年社会经济状况的交互作用会使个体做出两种截然不同的规避风险的行为, 来自贫困家庭的个体在感知到死亡威胁的信息时, 会采取多样化的选择以分散风险, 而来自富裕家庭的个体则会采取保守的行为方式, 通过在有限的事物上增大投资以减少风险。
2.3.3 生物因素
除了环境因素、童年经历外, 近期的研究发现生物因素也会影响生命史策略。目前发现的影响生命史策略的生物因素主要是睾丸酮水平 (Hormone testosterone)。Rosvall (2013)的研究结果显示, 个体的睾丸酮水平对于个体在求偶和养育间的权衡起到了决定性的作用, 雌性树燕被注射睾丸酮后会表现出攻击性增高, 孵化时体温降低使孵化成功率也会降低的结果, 表明睾丸酮会通过损害养育行为来影响生命史策略的发展。
3 生命史策略的测量
3.1 直接测量
个体的生命史策略是可以被直接测量的, 最常用的生命史量表是《亚利桑那生命史量表》 (Arizona Life History Battery; ALHB)及其分量表 Mini-K 量表(Figueredo et al., 2005)。ALHB 是一组测量与生命史策略相关的认知和行为指标的量表, 共有 199 个条目, 包括 Mini-K 分量表, 洞察、计划和控制力分量表, 父/母亲关系质量分量表、家庭/朋友联系分量表, 家庭/朋友支持分量表, 亲密关系经验分量表, 利他行为分量表以及信仰分量表。这些自陈式的心理指标可以从各方面衡量生命史策略, 分数越高, 表明个体越倾向于慢的生命史策略。此外, 由于 Mini-K 分量表的会聚效度最高, 其内部一致性系数和重测信度都达到了 0.7 以上, 因此为了方便施测, 研究者往往将 Mini-K 单独抽出来替代整个 ALHB 以减少被试的任务量(Figueredo et al., 2005)。
除了 ALHB 及其分量表 Mini-K 以外, 可以直接测量生命史策略的量表还有 High-K Strategy 量表(HKSS, Giosan, 2006)。与 Mini-K 不同, Mini-K 测量的是行为, 而 HKSS 则更多关注与慢生活史策略相关的一系列特质, 包括个人的生理状态及人际吸引力(Dunkel & Decker, 2010), 是否具有良好的健康状况、对环境安全/稳定性的知觉, 以及积极的自我概念/社会成就等(Olderbak, Gladden, Wolf, & Figueredo, 2014)。该量表由 23~26 个条目组成, 并且具有良好的结构效度和内部一致性系数(α = 0.92)。
3.2 间接测量
除了可以用量表直接测量个体的生命史策略, 研究者还可以通过影响生命史策略的环境因素、童年经验来间接测量生命史策略。
具体来说, 可以反映当下环境恶劣性的客观指标有:当下社会经济地位(收入水平、受教育程度)、当地犯罪率、当地疾病–死亡发生率以及预期寿命; 而其对应的童年经验为童年时期的社会经济地位(Griskevicius, Delton et al., 2011; Ellis et al., 2009; Griskevicius, Tybur et al., 2011; Frankenhuis et al., 2013; Simpson, Griskevicius, Kuo, Sung, & Collins, 2012)。
反映当下环境恶劣性的主观指标则包括:个体对生活环境的主观估计和感知, 对邻居暴力行为与犯罪状况的主观评定, 以及对亲属疾病或死亡情况的回忆; 相对应的童年经验包括对童年社会经济地位的主观评定, 以及对童年家庭环境冲突与暴力事件的主观评定 (Ellis et al., 2009; Griskevicius et al., 2013; White et al., 2013; McCullough, Pedersen, Schroder, Tabak, & Carver, 2013)。
反映当下环境不稳定性的测量指标主要是当地疾病–死亡发生率的波动情况(Ellis et al., 2009), 而对童年生活环境不稳定性的测量则可以通过父母生活压力的三种来源——失业、搬家与离异经历来测定(Simpson et al., 2012; Frankenhuis et al., 2013)。
除此之外, 最新关于生命史策略的研究还试图通过可靠的生物学指标进行测定, 其中一项有效的指标就是个体的氧化应激水平 (Oxidative Stress)。因为当有机体遭受各种有害刺激时, 体内的高活性分子(如活性氧自由基和活性氮自由基) 会产生过多, 使氧化系统和抗氧化系统失衡, 并最终导致组织损伤。因此, 个体较高的氧化应激水平表明他可能长期暴露于不良的、充满压力及低社会经济地位的环境中(Gangestad, Merriman, & Thompson, 2010)。在研究中, Griskevicius 等人用氧化应激水平的代表性标志——8-羟化脱氧鸟苷(8-OHdG)来间接测量个体的社会经济地位, 结果发现, 对于高氧化应激水平的个体(即低社会经济地位), 经济衰退的线索可以促使人们采取更快的生命史策略 , 比如做出更多的冒险行为 (Griskevicius et al., 2013)。
4 生命史策略与道德行为
通过上述对生命史理论的回顾, 研究者发现目前关于生命史理论的大多数研究都是针对个体发展的主题, 但是生命史策略不止与个体的成长轨迹有关, 也与个体当下所处的环境联系紧密 (Griskevicius, Delton et al., 2011)。快、慢生命史策略不仅可以体现在性成熟、生育等发展命题上, 还可以体现在个体的社会行为上。近期越来越多的生命史的研究开始逐渐关注到社会心理学领域, 以探究生命史与个体社会心理和行为之间的关系。比如有研究开始关注生命史与种族歧视 (Figueredo, Gladden, & Black, 2012; Gibbons et al., 2012; Neuberg & Sng, 2013)、社会敌意与敌对行为(Wenner, Bianchi, Figueredo, Rushton, & Jacobs, 2013)、投机行为(Ellis et al., 2012; Griskevicius et al., 2013; Griskevicius, Tybur et al., 2011; White et al., 2013) 以及社会信任等社会心理学变量 (Petersen & Aarøe, 2015)之间的关系。此外, 不仅在内容上有所侧重, 社会心理学中常用的研究方法也逐渐被扩展至生命史的研究中。具体来说, 以往的生命史研究主要依靠大量流调中自陈式报告的方法来探究变量之间的相关关系, 而社会心理学视角下的生命史研究逐渐开始采用情境测验、实验室行为观察、现场实验以及生态瞬时评估技术 (Sherman, Figueredo, & Funder, 2013; Laran & Salerno, 2013; White et al., 2013)这类社会心理学中常用的技术以使研究在方法上更加严谨并且具有可操作性。
因此, 本文就以亲社会行为和不道德/暴力犯罪行为为例, 总结已有文献中生命史策略对道德行为的关系以及不同的生活经历对其产生的影响, 并且提出可能解释这一影响的中介变量。本文不仅期望将生命史策略的范畴扩展至道德领域, 更期望为心理学家们在探索社会心理学中的其他问题时提供一个新的角度。
4.1 在亲社会行为上的体现
根据生命史理论, 生命史策略的不同会反映出个体不同的亲社会行为倾向, 快生命史策略的个体在感知到环境恶劣时可能做出更多的自利行为(如在公共资源困境游戏中更多地从公共资源中获取利益)、更少的亲社会行为(如更少地帮助他人), 并且与这些行为相关的人格因素(如自尊)也可能与不同的生命史策略有关。上述推理已经得到了部分相关研究的支持。
以往的调查研究已经发现慢生命史策略与道德直觉呈正相关(Gladden, Welch, Figueredo, & Jacobs, 2009)、与敌对态度/行为呈负相关(Wenner et al., 2013)。相比于快生命史策略冲动性、强烈的性驱动、缺乏强烈的情感依恋等特点, 持有慢生命史策略的个体在与环境的互动中认知成分更多, 思考更深思熟虑, 考虑的更长远, 他们更能抑制情绪冲动, 规范自己的行为(Figueredo et al., 2011), 因此不论是对内群体还是对其他人都更友善, 在社交中表现的更无私, 更倾向于互惠的、亲社会的行为策略(Figueredo et al., 2011)。已有研究证实, 在社交任务中, 慢生命史策略的个体往往表现出更多的社会赞许行为以及亲社会、合作行为(Figueredo & Rushton, 2009; Sherman et al., 2013), 甚至可以减少种族主义的发生(Figueredo et al., 2011)。相反, 恶劣的环境可以限制人们的群际融合, 已有研究发现当给白人被试呈现暗示环境恶劣的线索时, 他们更不愿意将黑白混血的个体归为内群体, 而当暗示环境良好的线索出现时, 这一效应就消失了(Rodeheffer, Hill, & Lord, 2012)。
不仅如此, 研究还发现慢生命史策略的个体通常对社会规范更敏感, 更遵守社会规范(Figueredo et al., 2011)。Figueredo 等人(2011)采用元分析的方法, 整合了多篇文章的数据, 发现 Mini-K 量表分数越高的个体, 即更偏向慢生命史策略的个体, 在一般人格因子、共赢社交策略、自我评价得分更高, 在敌对策略上的得分更低。其中一般人格因子分数越高, 意味着个体具有更高的开放性、责任心、外向性、宜人性, 以及更低的神经质; 共赢社交策略分数越高意味着个体更可能同时考虑自己和他人的利益, 做出共赢的决策; 自我评价的分数越高则意味着个体具有更高的自尊心、价值感; 在敌对社交策略上的分数越高意味着个体更可能为了自身利益损害他人利益。
研究者认为, 生命史策略之所以可以体现在个体亲社会行为上有以下两个可能的原因。首先, 慢生命史策略的个体更遵守社会规范可能是由于在相对稳定的环境中个体采取慢生命史策略更具有社会适应意义, 慢生命史策略本身就与环境的相对稳定和可预期相匹配, 因此在这样的环境下遵守社会规范通常更有利。并且由于人具有主观能动性, 个体通常会选择与其一贯行为方式相适应的环境, 以最大的发挥自身优势获得发展, 因此慢生命史策略的个体通常也会主动选择较为稳定、未来可预期这种与他们的策略相一致的环境, 以发挥慢生命史策略的优势。
其次, 生命史策略之所以与个体的亲社会行为有如此关系, 还可能是由于个体在社会交往中对人际关系的不同划分所导致的。具体来说, 以共享关系 (Communal relationship) 和交换关系 (Exchange relationship)这个社会心理学领域对人际关系比较常用的划分为例(Clark & Mills, 2011; Clark & Mils, 1993), 前者是指个体在关系中会更多地关注他人的需求, 而后者是指个体更多地关注自身需求(Miller et al., 2014)。由于快生命史策略的个体采取的是一种迅速满足个体需求、完成生命命题的策略, 那么他们相对于慢生命史策略的个体更可能将人际关系看成一种交换, 更看重自己的利益和对方可能带来的好处, 而较少关注对方的需求, 因此比慢生命史策略的个体做出更少亲社会行为。
此外, 对于另一些公益性的、有利于未来的亲社会行为, 比如环境保护问题, 进化心理学家们认为这些问题部分是由于人们更看重当下的满足, 而不是遥不可及的未来需要所产生的。如果个体当下生活于不稳定或极端的环境, 那么他们将采取一种较快的生命史策略, 在行为决策时往往更冲动, 更不关心不可预测的未来(Griskevicius, Cantú, & van Vugt, 2012), 因此也将表现出较少的公益性行为。
4.2 在不道德和犯罪行为上的体现
已有研究发现快生命史策略与个体的违规行为成正相关 (Bogaert & Rushton, 1989; Wenner et al., 2013), 而成长于高压力环境(比如贫穷、歧视、家庭暴力等)的儿童相比其他儿童在发展过程中更可能产生一些问题行为(Ellis et al., 2012)。具体来说, 如果父母对儿童漠不关心、将儿童置于家庭冲突不断的环境时, 儿童则会快速的发展和成熟, 这会影响男孩对道德的认识(如诚信), 他们可能会在日后的人际交往中做出剥削他人、报复他人的行为(McCullough et al., 2013)。即便控制了个体的经济地位、家庭结构等因素, 对未来不稳定性的感知仍可以有效预测个体的违规行为 (Caldwell, Wiebe, & Cleveland, 2006), 这可能是因为当青少年当下所处的环境和掌握的资源不能预测未来的良好发展, 或使之缺少竞争优势时, 参与冒险行为可以使他们获得额外的收益 (Wilson & Daly, 1985; Daly, Wilson, & Vasdev, 2001)。当然, 从进化心理学的角度来看, 冒险行为在某些情况下(极端和不稳定的环境)是具有相当高的适应意义的, 可能是个体当下最有利于自身的选择(Ellis et al., 2012)。
除了冒险和违规行为, 童年不稳定的成长环境还会增加个体日后的攻击性行为(Copping & Campbell, 2015)。Bjorklund 与 Hawley (2014)的研究发现, 早期不良的环境因素会促使个体在未来形成长期的攻击性行为模式, 比如在家暴环境中成长的儿童不论在家庭还是学校都比其他儿童表现出更多的攻击性(Dodge et al., 2008), 男孩可能更多的表现在肢体冲突上, 而女孩则更多表现在言语沟通与交往中(Cullerton-Sen et al., 2008)。以上证据都说明了快生命史策略可能和攻击行为有关。
近年来, 许多研究者都开始将生命史对个体行为的影响扩展到暴力犯罪行为上来, Copping 等人的研究发现个体感知到的不良环境线索(包括低预期寿命、无望的教育前景、逐年上升的失业率及离婚率、以及密集的人口)会增加年轻人的暴力行为 (Copping, Campbell, & Muncer, 2013); Copping 与 Campbell (2015)还认为个体早期不良的生活体验会使他们对未来的看法更悲观, 行为上更易不诚信和机会主义。不仅如此, van der Linden 等人还从人格角度对生命史策略和违法行为之间的关系进行研究 , 发现普遍人格因子 (General Factor of Personality)——除快、慢生命史策略外, 个体生命史策略的另一种表现形式—— 可以预测人们的犯罪行为, 即 GFP 得分更低的罪犯在行为上更加暴力, 犯罪记录也更多(van der Linden, Dunkel, Beaver, & Louwen, 2015)。这类研究结果也被实验室研究所证实, Dunkel, Mathes 和 Beaver (2013)发现, 操纵个体的预期寿命(Life expectancy)可以显著引起犯罪意图的改变。
然而快生命史策略的个体更易做出不道德行为这个假设, 并不适用于所有类型的行为。就以犯罪行为为例, 除了冲动犯罪这种来源于快生命史策略和低自控能力的犯罪行为以外, 还有一类是基于竞争优势的精英犯罪(Wiebe, 2012), 这类犯罪是个体经过深思熟虑的长期行为, 表现为自私地、毫无愧疚地掌控他人, 以自我为中心和不合理地高估自我价值。因而精英犯罪的特征与慢生命史策略更为相似, 但目前并没有研究证实这种推论, 到底快、慢生命史策略哪个更易产生精英犯罪, 目前还是一个待研究问题。
4.3 生命史影响道德行为的可能机制
目前绝大部分关于生命经验对个体行为策略影响的研究, 都是直接验证二者的因果关系, 但是这一影响的中介变量却很少触及。之前的研究仅涉及到控制感这一中介变量 , Mittal 和 Griskevicius (2014)的研究发现, 控制感在童年经验对个体冲动行为的影响中起到了中介作用, 他们的实验结果发现, 在面对不确定性时, 童年贫困的被试比童年富裕的被试具有更低的控制感, 而低控制感会进一步让他们做出冲动选择、在做任务时更不容易坚持等倾向, 但是如果在经历不确定性的同时给被试控制感, 两类被试的差异就消失了。该研究结果说明童年经验之所以会让个体形成快或者慢的生命史策略, 是因为它影响了控制感的形成。从这一结果带来灵感, 控制感或许可以解释为什么快生命史策略的个体会做出更多的不道德和暴力犯罪行为。
除控制感以外, 安全感与调节定向也可能成为生命史策略对个体道德行为产生影响的可能的中介变量。对安全感来说, 在前文理论综述中我们提到过, 在童年时期, 儿童可以通过父母投入于自身的关爱与资源的质量感受到外部环境中有关风险和机会的信息(Ellis et al., 2009)。因此童年生活富裕或是得到的关爱较多的孩子, 他们更可能形成对外部世界的积极认识, 获得更多的安全感, 而这种早期经历所塑造的安全感正可能是影响个体成年后面对环境变化而产生亲社会还是不道德行为的可能原因。而对调节定向来说 , Higgins (2014, 1998)在调节定向理论中将个体的动机分为促进(promotion)、预防(prevention)两类定向, 具体来说, 促进定向的个体在实现目标的过程中更关注收获(gain), 而预防定向的个体则更关注损失(loss)。由于快生命史策略的个体遭受过更多的资源匮乏、环境恶劣, 因此他们更可能产生一种“不会更差”的心理, 从而不会在意损失, 只关注收益, 即具有更强的促进定向动机, 这一动机或许可以解释为什么快生命史策略的个体更偏好冒险、冲动、违背社会规范的行为。——论文作者:彭芸爽 1 王 雪 2 吴 嵩 1 金盛华 1 孙荣芳 3
* 稍后学术顾问联系您




