
深海研究中的底栖有孔虫:回顾与展望
分类:农业论文
时间:2022-02-15
摘 要:底栖有孔虫在古环境研究中的应用先是用作古水深或水团的标志物。随着新技术的应用‚ 识别出了2种不同的底栖有孔虫微生境:外生种和内生种;认识到甚至深海底栖有孔虫‚也能对表层浮游生物勃发的季节性短暂事件作出响应‚因为沉降到海底的有机物质供养着底栖有孔虫。目前‚底栖有孔虫被广泛应用于估算海洋表层生产力和底层水团的含氧量。回顾了深海底栖有孔虫生态研究的历史和其在古海洋学中的应用‚并强调研究、应用中的新方法、新技术。中国已经加入了诸如 IODP 等深海研究计划‚有必要向我国学术界提供底栖有孔虫研究的新方向‚以资参考。
关 键 词:底栖有孔虫;深海研究;有机碳通量;含氧量;综合大洋钻探
底栖有孔虫是微体古生物学的起点。第一次世界大战前后‚美国科学家首先将这类微体化石作为地层年代的标志用于石油勘探‚大获成功‚从而建立了微体古生物学这门新学科。至于浮游有孔虫的地层意义‚要到20世纪50年代才被发现‚钙质超微化石的应用更晚。在现代的地质科学中‚微体化石首先是古海洋学、古环境研究的手段‚浮游有孔虫和钙质超微化石的应用比底栖有孔虫更为广泛。是不是底栖有孔虫已经“退居二线”‚在深海研究中不那么重要了呢?
本文从历史回顾入手‚分析底栖有孔虫研究及其应用的变化历程‚指出其在深海研究中不可替代的重要性。不是底栖有孔虫的研究价值发生了变化‚而是它所解的科学问题和研究的方法已经转移‚ 值得我国地学界、尤其是古海洋学界注意。
1 底栖有孔虫作为“标志种”
底栖有孔虫在早期的深海研究中‚和其它古生物化石应用于地层学或古生态学时作为“标志”物一样‚常常是作为深度或者水团的标志。
早在20世纪六七十年代以前‚已经发现了深海底栖有孔虫组合与深度相关‚虽然那时对其生态了解不多‚但普遍把底栖有孔虫用作古深度标志[1]‚ 认为“底栖有孔虫群是在岸外按水深呈带状分布” [2]。我国在研究海区底栖有孔虫生态分布的过程中‚也首先注意到与深度的关系。后来的研究表明‚底栖有孔虫的深度分布‚很大程度上是水团与水深的关系‚在不同深度处分布着不同的水团‚被不同水团占据的海底生活着不同属种组合的底栖有孔虫。因此‚在研究古深度时‚要求对古水体做综合性解释[3]‚不能用一个海区的深度资料直接去解释另一海区的古深度[4]。
与此同时‚底栖有孔虫的“标志种”也被广泛用于指示水团‚比如在大西洋深水区‚Epistominella umbonifera 指示南极底层水‚Epistominella exigua 和 Cibicidoides w uellerstorf i 指 示 北 大 西 洋 深 层水[5]。与水深相比‚水团代表的是多种环境因素的综合‚而不只是单一的环境因素[6]‚将底栖有孔虫应用于水团研究显然是一种进步。但无论水深或者水团‚底栖有孔虫直到80年代初还只是作为“标志种”在使用。尽管用虎红染色识别“活”有孔虫(其实是检查有无原生质)来研究深海底栖有孔虫生态的技术早在50年代就已经发明‚但由于采样和观测技术的限制‚这种生态研究在当时还只能是静态的、低分辨率的‚与今天以定量分析为基础、以追索机理为目标的古海洋学要求相比‚便显得不相适应。
2 深海底栖有孔虫微生境
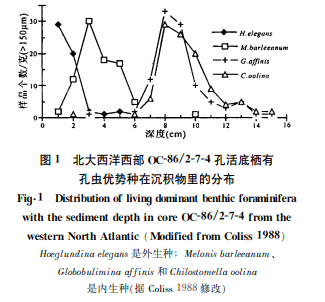
80年代起‚海上技术的发展为底栖有孔虫生态研究开辟了新阶段。美国 Corliss [7]把箱式取样器取上的 OC-86/2-7-4孔深海沉积柱状样切成1cm 厚的薄层‚在船上分别用虎红染色‚结果发现底栖有孔虫不仅分布在海底沉积层的表面‚还生活在沉积物之中‚垂向上呈层状分布(图1)‚证实了他以前根据据底栖有孔虫壳体的碳同位素差异推测某些底栖有孔虫可能生活在沉积物里的设想‚因而提出了底栖有孔虫微生境(microhabitat)的概念‚将底栖有孔虫区分为外(表)生种和内生种两大类(实际上还存在一类介于外生种和内生种之间的机会种)。
有关微生境的研究是底栖有孔虫生态学的一个重要进展。与一般生物的生境概念不同‚底栖有孔虫“微生境”涉及的是海水与沉积物界面上和界面以下沉积物里的垂向分布‚强调有孔虫活动的相对位置。在 Coliss [7]发现底栖有孔虫在海底沉积物里呈层状分布的同时‚他还发现了壳体形态与微生境的关系:具有旋卷型壳体的底栖有孔虫中‚表生种具有螺旋壳和双凸或平-凸的壳型‚便于在沉积物表面保持稳定;内生种往往具有平旋壳和浑圆的壳缘‚便于在沉积层内部移动。由于沉积层表层和内部含氧量不同‚表生种壳体的细孔可以只在背面发生‚而内生种的细孔则在壳面均匀分布‚以适应原生质与外界气体交换的需要。
不仅如此‚Corliss [8]在挪威海的研究结果表明‚ 底栖有孔虫的形态特征与深度分布具有相关性(图 2)‚而这种相关性实际上反映了微生境与深度的关系。壳体形态是锥形、圆柱型、扁平锥型、浑圆平旋型、圆球型或扁平椭圆型的内生种‚趋向于分布在 500~1000m 之间相对浅的海底;壳体形态是平-凸螺旋型、双凸螺旋型、粟状螺旋型和浑圆螺旋型的外生种‚趋向于分布在1500m 以下的海底;1000 ~1500m 则是二者之间的过渡‚200~500m 处可能因二者受生物扰动而含量基本相当。墨西哥湾的研究结果与挪威海的研究结果基本一致:1300m 以上内生种占主导地位‚2000m 以下主要是外生种‚1300~2000 m 是内生种和外生种之间的过渡[9]。微生境与深度的关系‚又取决于有机碳的通量:有机碳通量高的海底‚以内生种为主‚对应的海水深度较浅;有机碳通量低的海底‚水深较大‚以外生种为主‚也就是说‚有机碳通量控制着底栖有孔虫外生种和内生种的比例。譬如‚挪威海有机碳通量和微生境的变化以1000m 为界(图2)。
微生境的另一个方面是海底的氧化还原状况。表生种与内生种涉及两类不同的溶解氧:海底水体的溶解氧和海底沉积物孔隙水里的溶解氧。海底水体的溶解氧浓度是深部水团的一个重要性质‚对于更好地理解海洋环流历史、气候变化等具有重要意义。孔隙水里的溶解氧浓度对有孔虫的壳壁特征及其生活的微生境有重要影响‚内生种壳体只有比外生种壁孔密度大得多‚才能增强与外界的气体交换[7]。有关溶解氧对有孔虫影响的研究‚以 Kaiho [10]的工作最具代表性‚他建立了钙质底栖有孔虫氧浓度指数‚认为溶解氧浓度限制着底栖有孔虫的微生境:在氧化环境(oxic‚>1.5mL/L O2)里‚底栖有孔虫以外生种占优势‚具有壳体厚、壳径大(≥350 μm)的形态特征;在弱氧化环境(dysoxic‚0.1~0.3 mL/L O2)里‚底栖有孔虫组成主要是内生种‚具有壳体薄、壳径小(螺旋壳<200μm‚伸长扁平壳< 250μm)的特点;唯独在次氧化环境(suboxic‚0.3~ 1.5 mL/L O2)里‚外生种和内生种可以“和平共处”‚但也要以减小个体大小(<350μm)为代价。氧化环境里的外生种和弱氧化环境里的内生种生活在氧含量高(>2mL/L O2)的底层水下;而次氧化环境的有孔虫一般在低氧底层水中常见、但在高氧底层水环境里和接近缺氧环境里也可以生存。Kaiho 利用深海底栖有孔虫来研究溶解氧含量是深部水团性质研究中的一个典范。
微生境的研究‚导致了控制因素之争[11]:部分学者认为含氧量是控制因素‚而另一部分学者则认为营养物供应是最重要的控制因素。近年来对海洋表层生产力与底栖有孔虫关系的研究‚为回答这个问题提供了重要依据。
3 营养物质供应与底栖有孔虫
深海底栖有孔虫的食物来源与其生存环境密切相关。深海底栖有孔虫大致生活在两类环境里:全年低温、高水压、完全黑暗的一般深海环境和有深海热液、冷水渗透等的特殊环境[12]。在一般深海环境里‚海洋表层生产力是底栖有孔虫的营养物质提供者‚营养物质供应往往是季节性变化的;而在特殊环境里‚营养物质供应是非季节性的‚食物来源也很多样‚与前者相比‚人们对其认识还很肤浅。
3.1 季节性食物供应对底栖有孔虫的影响
海洋表层生产力是底栖有孔虫营养物质的主要提供者‚从海洋表面沉降至海底的营养物质维持着一般深海环境里的底栖有孔虫种群的生存和繁衍‚ 沉到海底的有机碳越多‚有孔虫丰度也就越高。曾经推算‚每1mg 的有机碳降到海底‚就可以有一个壳径>150μm 的底栖有孔虫生成(因此提出用底栖有孔虫的堆积速率估算古生产力[13])。由于海面藻类勃发后沉降到海底的植物碎屑先呈浮浆(fluff)状存在‚这类浮浆就为追踪底栖生物对表层生产力事件的响应提供了材料。果然‚80年代中期‚英国 Gooday 在北大西洋4000m 左右水深处采集到海底浮浆‚从中发现了大量的底栖有孔虫活个体‚其中以 Epistominella exigua 等3个种占据优势[14]。这项发现改变了深海底栖有孔虫对海洋表层环境反映不灵敏的偏见‚证明活有孔虫群在海洋底部也能响应季节性变化和表层海水的事件[15]。
更加系统的研究来自日本。Kitazato 等[12]在东京以南的相模湾千余米的深水处‚进行了4年(1994-1998)的定点观测‚采用遥测遥感(海洋生产力)、沉积捕集器(沉降颗粒)、海底录像(雾状层[nepheloid layer]观测)和多管取样(沉积物及表面的浮浆)等多种手段‚发现海面藻类勃发(图3A)后‚不仅沉积通量增大(图3B)‚而且海底的雾状层变浓(图 3C)‚浮浆层增厚(图3D)‚沉积物氧化层减薄(图3E)‚ 活底栖有孔虫也相应增多(图3F)‚生动地展现了表层生产力驱使底栖有孔虫繁盛的过程。
90年代以来‚浮游生物勃发事件对底栖有孔虫的影响‚已经引起学术界的密切关注‚使得有关表层生产力或者有机碳通量与底栖有孔虫属种组合的关系趋于明朗。在生物勃发期间‚沉降至海底的有机碳总量占年有机物总量的大部分[16]‚并且它们多不稳定‚降解快。底栖有孔虫适应这种环境‚特化出了短期迅速繁殖生长和长期忍受饥饿的能力‚从而产生出随有机碳通量变化的属种组合:表生种和胶结质浅内生种是低有机碳通量海底的特征组合;而在高有机碳通量的海底‚有机会种(也叫暂时性表生种)和内生种组合(图4)。不仅如此‚不同海区的表层生产力季节性变化往往很不一样‚其强弱和有机物质向海底间歇式沉积的频率也会影响底栖有孔虫属种组合[17]。
有关有机碳通量与底栖有孔虫组合关系的新认识‚是对 Coliss 等[8]发现的在不同水深处有机碳通量和微生境关系的进一步发展‚也为有孔虫组合的深度分布提供了解释:不同深度海区的生产力和有机碳通量不同‚导致了底栖有孔虫属种分布的不同。以地中海为例‚表层生产力由西向东递减‚因此同一深度的底栖有孔虫组合也呈东西向变化;而同一种的深度分布界限也随着有机碳通量的不同而变化[20]‚譬如外生种 Gyroidina orbicularis 生活在较深海底‚占全群落10%以上的高值区上限‚在西地中海为1500m‚向东则上升到800~600m。
3.2 非季节性食物供应对底栖有孔虫的影响
在深入研究表层生产力季节性变化对深海底栖有孔虫影响的同时‚非季节性食物输入对深海底栖有孔虫的影响也引起了人们的注意。由于非季节性食物输入的种类较多且较复杂‚目前对其和底栖有孔虫的关系还了解不多‚像鲸尸、无脊椎动物、木材等营养物质向海底的输入没有规律‚也不可预测‚在大多数地方具有局部性‚它们对底栖有孔虫的影响与自身大小和性质有关;而在低强度浊流频繁发生的地方‚底栖有孔虫能够依靠有机物质持续生存;在有深海热液渗透的海底‚有孔虫可能是当地动物群的重要组分‚其密度、多样性、组成多不同于非热液区动物群[16]。一般说来‚适应非季节性营养供应的有孔虫往往是那些能适应低氧和(或)高生产力环境的物种‚它们响应有机物输入的方式可能因有机物本身的性质不同和输入的规模和时间尺度不同而有差别[18]。
总之‚深海研究中的底栖有孔虫已由当初的古深度和水团的“标志种”‚变成了能够揭示溶解氧含量和表层生产力变化(也能反映有机物非季节性变化)的“要员”‚成为一身兼多职的“多面手”。这固然说明其应用范围变宽‚但也标志着对底栖有孔虫研究程度的深入。实际上‚海水深度(或水团)、有机碳通量、溶解氧含量之间存在某种关联性。在分布着不同水团的海水深度处‚有机碳通量往往也不一样‚ 而有机碳通量与溶解氧含量通常呈现负相关性。它们之间的关系实际上是海水深度与微生境之间的关系。
回到微生境控制因素的题目上来‚食物供应和氧浓度是微生境的两大控制因素(图5)。在缺营养和富营养的环境里‚只有外生种和浅内生种能够生存‚因为在缺营养的环境里‚沉降下来的有机物大部分在沉积物表面被消耗、氧化‚仅有少量难溶的有机物经过生物扰动等作用能进入浅内生境里‚食物供应限制着有孔虫的生存和繁殖;而在富营养环境里‚ 氧浓度是限制因子(氧含量低于0.5mL/L 时[18])‚ 大量的有机物被输送到海底‚消耗大量溶解氧‚深的内生境因缺氧而无深内生种生存。只有在中等营养环境里‚各种微生境的有孔虫才能共同生存‚因为除了部分有机物被外生底栖有孔虫消耗、氧化外‚还有部分有机物质进入深的内生境里‚在这种情况下‚食物供应和氧浓度共同限制着底栖有孔虫在沉积物里的最深分布。食物供应太少或者太多‚对很多底栖有孔虫都不利‚食物过于贫乏会导致营养供应不足‚ 过于丰富会导致沉积物处于还原环境‚这两种极端均导致底栖有孔虫以特殊或单一的外生种为主‚甚至导致底栖有孔虫消失。
4 展望新世纪深海底栖有孔虫研究
纵观深海底栖有孔虫研究‚可以发现‚海底有机碳通量和深部水团这两大因素控制着底栖有孔虫的分布。如果说底栖有孔虫在海底沉积物里的垂向分布受营养供应和氧浓度的控制‚那么‚其在洋底横向上的分布则受当地水团所控制。海水沉积物和海水界面处和界面以下的沉积物里的氧浓度‚既和营养供应有关‚又和水团本身的氧浓度有关。假如海底有机碳通量不变‚被性质不同的水团所覆盖的海底微生境的“主人”也会很不相同。因此‚我们研究底栖有孔虫‚总的来说主要可做两件基本方面的事情:一是利用底栖有孔虫研究深部水团的变化规律‚二是利用底栖有孔虫研究海洋表层初级生产力。前者反映地球表层系统中的水循环‚后者涉及碳循环‚都是地球系统科学中的基本问题。
进入新世纪‚传统的描述性的微体古生物学已经转入新一代的古环境研究‚不同之处在于定量分析的方法、追索机理的目标和多学科的途径。作为深海底栖生物中最为常见、又能保留化石的门类‚底栖有孔虫正成为深海古海洋学研究的“主角”‚展现着“第二个春天”的前景。与此相应‚研究方法、观测技术的创新已经成为时代的要求‚如为求得更完整的生态指标‚在冷藏室中模拟海底温度饲养深海底栖有孔虫、采用荧光等新技术识别活有孔虫等[21]‚ 都是有益的尝试。
现场的实时观测‚将是新世纪深海研究的重要新方向‚底栖有孔虫为探测深海海底的过程提供了灵敏的检测器。如墨西哥湾北部500~600m 水深处烃类泄出口发现的底栖有孔虫‚说明在总体处于还原环境下的海底还可以有短暂间歇让有孔虫生存[22]。像日本相模湾式的定点观察‚从海面到海底的系统研究‚势必在新世纪的深海研究中进一步推广‚成为揭示海洋过程的新手段。
当前的古海洋学研究‚总体偏重海水的上层结构‚对于海洋深部的信息明显缺乏。底栖有孔虫是唯一从海岸到深海盆地无所不在而又大量出现、可以保存的生物门类‚是将古海洋学推进到整个海水系统的重要支撑。通过不同深水海底的底栖有孔虫取样研究‚可望取得中层水、深层水和底层水的全套信息‚必将有助于从根本上加深对地球系统的理解。
底栖有孔虫的钙质壳体‚广泛用于各种稳定同位素(如δ13C‚δ18O 等)和元素比值(如 Cd/Ca‚Mg/ Ca‚Sr/Ca)分析‚已经成为古海洋学的基本方法之一。可惜对这些有孔虫的生态特征了解有限‚以至于对其古环境解释基本上是单纯的矿物化学分析‚ 并不考虑控制这类壳体生长的有孔虫生态学特征。因此新世纪的一大任务是扩展深海有孔虫的生态研究‚只有在了解其微生境的特征后‚才可能正确解释其壳体的化学成分[23]。
和其它门类的化石一样‚底栖有孔虫的壳体也经历着埋藏学的筛选:胶结壳的分解和钙质壳的溶解‚导致埋藏群、化石群与生物群之间的差异。因此‚埋藏学的研究是根据化石群再造古生态环境的必要环节‚加强底栖有孔虫埋藏学研究‚势必有利于古环境研究。
我国的古生物学研究具有光荣的历史传统和长期的科学积累。作为古海洋学、古环境研究手段的底栖有孔虫研究起步较晚‚但近20年已经取得了长足的进步。中国自1998年加入国际大洋钻探以来‚ 首先是以微体古生物学为基础的古海洋学研究进入了国际学术前沿‚开创了我国深海研究的新局面。 2003年10月‚国际综合大洋钻探(IODP)计划启动‚我国以加倍的投入继续成为其参与成员国‚而古环境研究是 IODP 的三大科学主题之一[24]‚也是我国可以发挥作用的首选领域。近年来‚我国的深海底栖有孔虫研究已经走向国际(如文献25)‚在新的国际计划中应当将属种组合的研究与地球化学的多种指标相结合‚将室内镜下分析与海上现场观测、试验和数值模拟相结合‚争取在更高的层次上参与国际学术合作和竞争。——论文作者:张江勇‚汪品先
* 稍后学术顾问联系您




